What is the Membrrane That Forms a Continuous Cheet
The lipid bilayer has been firmly established as the universal basis for cell-membrane structure. It is easily seen by electron microscopy, although specialized techniques, such as x-ray diffraction and freeze-fracture electron microscopy, are needed to reveal the details of its organization. The bilayer structure is attributable to the special properties of the lipid molecules, which cause them to assemble spontaneously into bilayers even under simple artificial conditions.
Membrane Lipids Are Amphipathic Molecules, Most of which Spontaneously Form Bilayers
Lipid—that is, fatty—molecules constitute about 50% of the mass of most animal cell membranes, nearly all of the remainder being protein. There are approximately 5 × 106 lipid molecules in a 1 μm × 1 μm area of lipid bilayer, or about 109 lipid molecules in the plasma membrane of a small animal cell. All of the lipid molecules in cell membranes are amphipathic (or amphiphilic)—that is, they have a hydrophilic ("water-loving") or polar end and a hydrophobic ("water-fearing") or nonpolar end.
The most abundant membrane lipids are the phospholipids. These have a polar head group and two hydrophobic hydrocarbon tails. The tails are usually fatty acids, and they can differ in length (they normally contain between 14 and 24 carbon atoms). One tail usually has one or more cis-double bonds (i.e., it is unsaturated), while the other tail does not (i.e., it is saturated). As shown in Figure 10-2, each double bond creates a small kink in the tail. Differences in the length and saturation of the fatty acid tails are important because they influence the ability of phospholipid molecules to pack against one another, thereby affecting the fluidity of the membrane (discussed below).
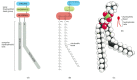
Figure 10-2
The parts of a phospholipid molecule. This example is phosphatidylcholine, represented (A) schematically, (B) by a formula, (C) as a space-filling model, and (D) as a symbol. The kink resulting from the cis-double bond is exaggerated for emphasis.
It is the shape and amphipathic nature of the lipid molecules that cause them to form bilayers spontaneously in aqueous environments. As discussed in Chapter 2, hydrophilic molecules dissolve readily in water because they contain charged groups or uncharged polar groups that can form either favorable electrostatic interactions or hydrogen bonds with water molecules. Hydrophobic molecules, by contrast, are insoluble in water because all, or almost all, of their atoms are uncharged and nonpolar and therefore cannot form energetically favorable interactions with water molecules. If dispersed in water, they force the adjacent water molecules to reorganize into icelike cages that surround the hydrophobic molecule (Figure 10-3). Because these cage structures are more ordered than the surrounding water, their formation increases the free energy. This free energy cost is minimized, however, if the hydrophobic molecules (or the hydrophobic portions of amphipathic molecules) cluster together so that the smallest number of water molecules is affected.

Figure 10-3
How hydrophilic and hydrophobic molecules interact differently with water. (A) Because acetone is polar, it can form favorable electrostatic interactions with water molecules, which are also polar. Thus, acetone readily dissolves in water. (B) By contrast, (more...)
For the above reason, lipid molecules spontaneously aggregate to bury their hydrophobic tails in the interior and expose their hydrophilic heads to water. Depending on their shape, they can do this in either of two ways: they can form spherical micelles, with the tails inward, or they can form bimolecular sheets, or bilayers, with the hydrophobic tails sandwiched between the hydrophilic head groups (Figure 10-4).

Figure 10-4
Packing arrangements of lipid molecules in an aqueous environment. (A) Wedge-shaped lipid molecules (above) form micelles, whereas cylinder-shaped phospholipid molecules (below) form bilayers. (B) A lipid micelle and a lipid bilayer seen in cross section. (more...)
Being cylindrical, phospholipid molecules spontaneously form bilayers in aqueous environments. In this energetically most-favorable arrangement, the hydrophilic heads face the water at each surface of the bilayer, and the hydrophobic tails are shielded from the water in the interior. The same forces that drive phospholipids to form bilayers also provide a self-healing property. A small tear in the bilayer creates a free edge with water; because this is energetically unfavorable, the lipids spontaneously rearrange to eliminate the free edge. (In eucaryotic plasma membranes, larger tears are repaired by the fusion of intracellular vesicles.) The prohibition against free edges has a profound consequence: the only way for a bilayer to avoid having edges is by closing in on itself and forming a sealed compartment (Figure 10-5). This remarkable behavior, fundamental to the creation of a living cell, follows directly from the shape and amphipathic nature of the phospholipid molecule.

Figure 10-5
The spontaneous closure of a phospholipid bilayer to form a sealed compartment. The closed structure is stable because it avoids the exposure of the hydrophobic hydrocarbon tails to water, which would be energetically unfavorable.
A lipid bilayer has other characteristics beside its self-sealing properties that make it an ideal structure for cell membranes. One of the most important of these is its fluidity, which is crucial to many membrane functions.
The Lipid Bilayer Is a Two-dimensional Fluid
It was only around 1970 that researchers first recognized that individual lipid molecules are able to diffuse freely within lipid bilayers. The initial demonstration came from studies of synthetic lipid bilayers. Two types of preparations have been very useful in such studies: (1) bilayers made in the form of spherical vesicles, called liposomes, which can vary in size from about 25 nm to 1 μm in diameter depending on how they are produced (Figure 10-6); and (2) planar bilayers, called black membranes, formed across a hole in a partition between two aqueous compartments (Figure 10-7).
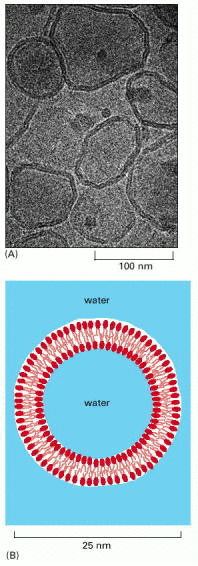
Figure 10-6
(A) An electron micrograph of unfixed, unstained phospholipid vesicles—liposomes—in water rapidly frozen to liquid nitrogen temperature. The bilayer structure of the liposomes is readily apparent. (B) A drawing of a small spherical liposome (more...)

Figure 10-7
A cross-sectional view of a black membrane, a synthetic lipid bilayer. This planar bilayer appears black when it forms across a small hole in a partition separating two aqueous compartments. Black membranes are used to measure the permeability properties (more...)
Various techniques have been used to measure the motion of individual lipid molecules and their different parts. One can construct a lipid molecule, for example, whose polar head group carries a "spin label," such as a nitroxyl group (>N-O); this contains an unpaired electron whose spin creates a paramagnetic signal that can be detected by electron spin resonance (ESR) spectroscopy. (The principles of this technique are similar to those of nuclear magnetic resonance, discussed in Chapter 8.) The motion and orientation of a spin-labeled lipid in a bilayer can be deduced from the ESR spectrum. Such studies show that phospholipid molecules in synthetic bilayers very rarely migrate from the monolayer (also called a leaflet) on one side to that on the other. This process, known as "flip-flop," occurs less than once a month for any individual molecule. In contrast, lipid molecules readily exchange places with their neighbors within a monolayer (~107 times per second). This gives rise to a rapid lateral diffusion, with a diffusion coefficient (D) of about 10-8 cm2/sec, which means that an average lipid molecule diffuses the length of a large bacterial cell (~2 μm) in about 1 second. These studies have also shown that individual lipid molecules rotate very rapidly about their long axis and that their hydrocarbon chains are flexible (Figure 10-8).

Figure 10-8
Phospholipid mobility. The types of movement possible for phospholipid molecules in a lipid bilayer.
Similar studies have been performed with labeled lipid molecules in isolated biological membranes and in living cells. The results are generally the same as for synthetic bilayers, and they demonstrate that the lipid component of a biological membrane is a two-dimensional liquid in which the constituent molecules are free to move laterally. As in synthetic bilayers, individual phospholipid molecules are normally confined to their own monolayer. This confinement creates a problem for their synthesis. Phospholipid molecules are made in only one monolayer of a membrane, mainly in the cytosolic monolayer of the endoplasmic reticulum (ER) membrane. If none of these newly made molecules could migrate reasonably promptly to the noncytosolic monolayer, new lipid bilayer could not be made. The problem is solved by a special class of membrane-bound enzymes called phospholipid translocators, which catalyze the rapid flip-flop of phospholipids from one monolayer to the other, as discussed in Chapter 12.
The Fluidity of a Lipid Bilayer Depends on Its Composition
The fluidity of cell membranes has to be precisely regulated. Certain membrane transport processes and enzyme activities, for example, cease when the bilayer viscosity is experimentally increased beyond a threshold level.
The fluidity of a lipid bilayer depends on both its composition and its temperature, as is readily demonstrated in studies of synthetic bilayers. A synthetic bilayer made from a single type of phospholipid changes from a liquid state to a two-dimensional rigid crystalline (or gel) state at a characteristic freezing point. This change of state is called a phase transition, and the temperature at which it occurs is lower (that is, the membrane becomes more difficult to freeze) if the hydrocarbon chains are short or have double bonds. A shorter chain length reduces the tendency of the hydrocarbon tails to interact with one another, and cis-double bonds produce kinks in the hydrocarbon chains that make them more difficult to pack together, so that the membrane remains fluid at lower temperatures (Figure 10-9). Bacteria, yeasts, and other organisms whose temperature fluctuates with that of their environment adjust the fatty acid composition of their membrane lipids to maintain a relatively constant fluidity. As the temperature falls, for instance, fatty acids with more cis-double bonds are synthesized, so the decrease in bilayer fluidity that would otherwise result from the drop in temperature is avoided.

Figure 10-9
The influence of cis-double bonds in hydrocarbon chains. The double bonds make it more difficult to pack the chains together, thereby making the lipid bilayer more difficult to freeze. In addition, because the fatty acid chains of unsaturated lipids (more...)
The lipid bilayer of many cell membranes is not composed exclusively of phospholipids, however; it often also contains cholesterol and glycolipids. Eucaryotic plasma membranes contain especially large amounts of cholesterol (Figure 10-10)—up to one molecule for every phospholipid molecule. The cholesterol molecules enhance the permeability-barrier properties of the lipid bilayer. They orient themselves in the bilayer with their hydroxyl groups close to the polar head groups of the phospholipid molecules. In this position, their rigid, platelike steroid rings interact with—and partly immobilize—those regions of the hydrocarbon chains closest to the polar head groups (Figure 10-11). By decreasing the mobility of the first few CH2 groups of the hydrocarbon chains of the phospholipid molecules, cholesterol makes the lipid bilayer less deformable in this region and thereby decreases the permeability of the bilayer to small water-soluble molecules. Although cholesterol tends to make lipid bilayers less fluid, at the high concentrations found in most eucaryotic plasma membranes, it also prevents the hydrocarbon chains from coming together and crystallizing. In this way, it inhibits possible phase transitions.
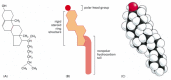
Figure 10-10
The structure of cholesterol. Cholesterol is represented (A) by a formula, (B) by a schematic drawing, and (C) as a space-filling model.

Figure 10-11
Cholesterol in a lipid bilayer. Schematic drawing of a cholesterol molecule interacting with two phospholipid molecules in one monolayer of a lipid bilayer.
The lipid compositions of several biological membranes are compared in Table 10-1. Bacterial plasma membranes are often composed of one main type of phospholipid and contain no cholesterol; their mechanical stability is enhanced by an overlying cell wall (see Figure 11-17). The plasma membranes of most eucaryotic cells, by contrast, are more varied, not only in containing large amounts of cholesterol, but also in containing a mixture of different phospholipids.
Table 10-1
Approximate Lipid Compositions of Different Cell Membranes.
Four major phospholipids predominate in the plasma membrane of many mammalian cells: phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine, and sphingomyelin. The structures of these molecules are shown in Figure 10-12. Note that only phosphatidylserine carries a net negative charge, the importance of which we discuss later; the other three are electrically neutral at physiological pH, carrying one positive and one negative charge. Together these four phospholipids constitute more than half the mass of lipid in most membranes (see Table 10-1). Other phospholipids, such as the inositol phospholipids, are present in smaller quantities but are functionally very important. The inositol phospholipids, for example, have a crucial role in cell signaling, as discussed in Chapter 15.
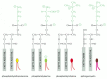
Figure 10-12
Four major phospholipids in mammalian plasma membranes. Note that different head groups are represented by different colors. All the lipid molecules shown are derived from glycerol except for sphingomyelin, which is derived from serine.
One might wonder why eucaryotic membranes contain such a variety of phospholipids, with head groups that differ in size, shape, and charge. One can begin to understand why if one thinks of the membrane lipids as constituting a two-dimensional solvent for the proteins in the membrane, just as water constitutes a three-dimensional solvent for proteins in an aqueous solution. Some membrane proteins can function only in the presence of specific phospholipid head groups, just as many enzymes in aqueous solution require a particular ion for activity. Moreover, some cytosolic enzymes bind to specific lipid head groups exposed on the cytosolic face of a membrane and are thus recruited to and concentrated at specific membrane sites.
The Plasma Membrane Contains Lipid Rafts That Are Enriched in Sphingolipids, Cholesterol, and Some Membrane Proteins
Most types of lipid molecules in cell membranes are randomly mixed in the lipid monolayer in which they reside. The van der Waals attractive forces between neighboring fatty acid tails are not selective enough to hold groups of molecules of this sort together. For some lipid molecules, however, such as the sphingolipids (discussed below), which tend to have long and saturated fatty hydrocarbon chains, the attractive forces can be just strong enough to hold the adjacent molecules together transiently in small microdomains. Such microdomains, or lipid rafts, can be thought of as transient phase separations in the fluid lipid bilayer where sphingolipids become concentrated.
The plasma membrane of animal cells is thought to contain many such tiny lipid rafts (~70 nm in diameter), which are rich in both sphingolipids and cholesterol. Because the hydrocarbon chains of the lipids concentrated there are longer and straighter than the fatty acid chains of most membrane lipids, the rafts are thicker than other parts of the bilayer (see Figure 10-9) and can better accommodate certain membrane proteins, which therefore tend to accumulate there (Figure 10-13). In this way, lipid rafts are thought to help organize such proteins—either concentrating them for transport in small vesicles or to enable the proteins to function together, as when they convert extracellular signals into intracellular ones (discussed in Chapter 15).
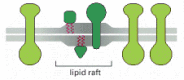
Figure 10-13
A lipid raft. Lipid rafts are small, specialized areas in membranes where some lipids (primarily sphingolipids and cholesterol) and proteins (green) are concentrated. Because the lipid bilayer is somewhat thicker in the rafts, certain membrane proteins (more...)
For the most part, lipid molecules in one monolayer of the bilayer move about independently of those in the other monolayer. In lipid rafts, however, the long hydrocarbon chains of the sphingolipids in one monolayer interact with those in the other monolayer. Thus, the two monolayers in a lipid raft communicate through their lipid tails.
The Asymmetry of the Lipid Bilayer Is Functionally Important
The lipid compositions of the two monolayers of the lipid bilayer in many membranes are strikingly different. In the human red blood cell membrane, for example, almost all of the lipid molecules that have choline—(CH3)3N+CH2CH2OH—in their head group (phosphatidylcholine and sphingomyelin) are in the outer monolayer, whereas almost all of the phospholipid molecules that contain a terminal primary amino group (phosphatidylethanolamine and phosphatidylserine) are in the inner monolayer (Figure 10-14). Because the negatively charged phosphatidylserine is located in the inner monolayer, there is a significant difference in charge between the two halves of the bilayer. We discuss in Chapter 12 how lipid asymmetry is generated and maintained by membrane-bound phospholipid translocators.

Figure 10-14
The asymmetrical distribution of phospholipids and glycolipids in the lipid bilayer of human red blood cells. The colors used for the phospholipid head groups are those introduced in Figure 10-12. In addition, glycolipids are drawn with hexagonal polar (more...)
Lipid asymmetry is functionally important. Many cytosolic proteins bind to specific lipid head groups found in the cytosolic monolayer of the lipid bilayer. The enzyme protein kinase C (PKC), for example, is activated in response to various extracellular signals. It binds to the cytosolic face of the plasma membrane, where phosphatidylserine is concentrated, and requires this negatively charged phospholipid for its activity.
In other cases, the lipid head group must first be modified so that proteinbinding sites are created at a particular time and place. Phosphatidylinositol, for instance, is a minor phospholipid that is concentrated in the cytosolic monolayer of cell membranes. A variety of lipid kinases can add phosphate groups at distinct positions in the inositol ring. The phosphorylated inositol phospholipids then act as binding sites that recruit specific proteins from the cytosol to the membrane. An important example of a lipid kinase is phosphatidylinositol kinase (PI 3-kinase), which is activated in response to extracellular signals and helps to recruit specific intracellular signaling proteins to the cytosolic face of the plasma membrane (Figure 10-15A). Similar lipid kinases phosphorylate inositol phospholipids in intracellular membranes and thereby help to recruit proteins that guide membrane transport.
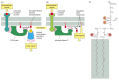
Figure 10-15
Some functions of membrane phospholipids in cell signaling. (A) Extracellular signals can activate PI 3-kinase, which phosphorylates inositol phospholipids in the plasma membrane. Various intracellular signaling molecules then bind to these phosphorylated (more...)
Phospholipids in the plasma membrane are used also in another way in the response to extracellular signals. The plasma membrane contains various phospholipases that are activated by extracellular signals to cleave specific phospholipid molecules, generating fragments of these molecules that act as short-lived intracellular mediators (Figure 10-15B). Phospholipase C, for example, cleaves an inositol phospholipid in the cytosolic monolayer of the plasma membrane to generate two fragments, one of which remains in the membrane and helps activate protein kinase C, while the other is released into the cytosol and stimulates the release of Ca2+ from the endoplasmic reticulum (see Figure 15-36).
Animals exploit the phospholipid asymmetry of their plasma membranes to distinguish between live and dead cells. When animal cells undergo programmed cell death, or apoptosis (discussed in Chapter 17), phosphatidylserine, which is normally confined to the cytosolic monolayer of the plasma membrane lipid bilayer, rapidly translocates to the extracellular monolayer. The phosphatidylserine exposed on the cell surface serves as a signal to induce neighboring cells, such as macrophages, to phagocytose the dead cell and digest it. The translocation of the phosphatidylserine in apoptotic cells occurs by two mechanisms:
- 1.
-
The phospholipid translocator that normally transports this lipid from the noncytosolic monolayer to the cytosolic monolayer is inactivated.
- 2.
-
A "scramblase" that transfers phospholipids nonspecifically in both directions between the two monolayers is activated.
Glycolipids Are Found on the Surface of All Plasma Membranes
The lipid molecules with the most extreme asymmetry in their membrane distribution are the sugar-containing lipid molecules called glycolipids. These intriguing molecules are found exclusively in the noncytosolic monolayer of the lipid bilayer, where they are thought to partition preferentially into lipid rafts. The glycolipids tend to self-associate, partly through hydrogen bonds between their sugars and partly through van der Waals forces between their long and mainly saturated hydrocarbon chains. The asymmetric distribution of glyco-lipids in the bilayer results from the addition of sugar groups to the lipid molecules in the lumen of the Golgi apparatus, which is topologically equivalent to the exterior of the cell (discussed in Chapter 12). In the plasma membrane, the sugar groups are exposed at the cell surface (see Figure 10-14), where they have important roles in interactions of the cell with its surroundings.
Glycolipids probably occur in all animal cell plasma membranes, where they generally constitute about 5% of the lipid molecules in the outer monolayer. They are also found in some intracellular membranes. The most complex of the glycolipids, the gangliosides, contain oligosaccharides with one or more sialic acid residues, which give gangliosides a net negative charge (Figure 10-16). More than 40 different gangliosides have been identified. They are most abundant in the plasma membrane of nerve cells, where gangliosides constitute 5–10% of the total lipid mass; they are also found in much smaller quantities in other cell types.
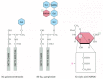
Figure 10-16
Glycolipid molecules. (A) Galactocerebroside is called a neutral glycolipid because the sugar that forms its head group is uncharged. (B) A ganglioside always contains one or more negatively charged sialic acid residues (also called N-acetylneuraminic (more...)
Hints as to what the functions of glycolipids might be come from their localization. In the plasma membrane of epithelial cells, for example, glycolipids are confined to the exposed apical surface, where they may help protect the membrane against the harsh conditions frequently found there (such as low pH and degradative enzymes). Charged glycolipids, such as gangliosides, may be important for their electrical effects: their presence alters the electrical field across the membrane and the concentrations of ions—especially Ca2+—at the membrane surface. Glycolipids are also thought to function in cell-recognition processes, in which membrane-bound carbohydrate-binding proteins (lectins) bind to the sugar groups on both glycolipids and glycoproteins in the process of cell-cell adhesion (discussed in Chapter 19). Surprisingly, however, mutant mice that are deficient in all of their complex gangliosides show no obvious abnormalities, although the males cannot transport testosterone normally in the testes and are consequently sterile.
Whatever their normal function, some glycolipids provide entry points for certain bacterial toxins. The ganglioside GM1 (see Figure 10-16), for example, acts as a cell-surface receptor for the bacterial toxin that causes the debilitating diarrhea of cholera. Cholera toxin binds to and enters only those cells that have GM1 on their surface, including intestinal epithelial cells. Its entry into a cell leads to a prolonged increase in the concentration of intracellular cyclic AMP (discussed in Chapter 15), which in turn causes a large efflux of Na+ and water into the intestine.
Summary
Biological membranes consist of a continuous double layer of lipid molecules in which membrane proteins are embedded. This lipid bilayer is fluid, with individual lipid molecules able to diffuse rapidly within their own monolayer. The membrane lipid molecules are amphipathic. The most numerous are the phospholipids. When placed in water they assemble spontaneously into bilayers, which form sealed compartments that reseal if torn.
There are three major classes of membrane lipid molecules—phospholipids, cholesterol, and glycolipids. The lipid compositions of the inner and outer monolayers are different, reflecting the different functions of the two faces of a cell membrane. Different mixtures of lipids are found in the membranes of cells of different types, as well as in the various membranes of a single eucaryotic cell. Some membrane-bound enzymes require specific lipid head groups in order to function. The head groups of some lipids form docking sites for specific cytosolic proteins. Some extracellular signals that act through membrane receptor proteins activate phospholipases that cleave selected phospholipid molecules in the plasma membrane, thereby generating fragments that act as intracellular signaling molecules.
Source: https://www.ncbi.nlm.nih.gov/books/NBK26871/
0 Response to "What is the Membrrane That Forms a Continuous Cheet"
Post a Comment